当社は埼玉大学発スタートアップとして2016年8月に設立されました。1985年頃から埼玉大学では、伏見譲博士らによって進化分子工学(Evolutionary Molecular Engineering)の研究が進められてきました。
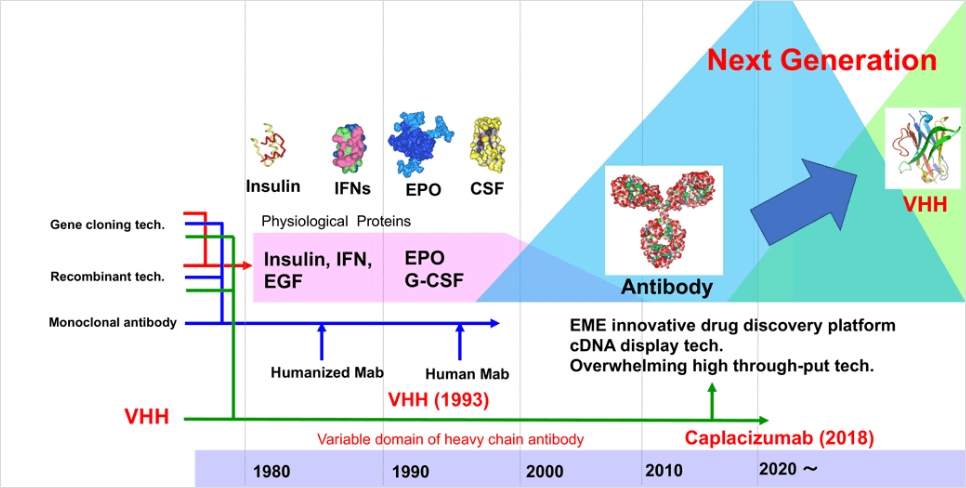
近年世界でも進化分子工学は注目されており、「ペプチドと抗体のファージディスプレイ法」を開発したGeorge P. Smith 博士が2018年度ノーベル化学賞を受賞しました。一方、伏見博士、当社代表取締役根本らによって無細胞翻訳系を用いたディスプレイ法であるin vitro virus (mRNAディスプレイ)法が、世界に先駆けて開発された歴史があります。しかし、mRNAディスプレイ法はファージディスプレイ法の約1万倍の効率を持つものの、安定性の面で難がありました。そこでmRNAディスプレイ方法の難点を解決した新たなcDNAディスプレイ法の開発が行われ、安定性の向上、質・量(=配列多様性)ともに既存のディスプレイ法を超える特徴を持つ遺伝子型-表現型分子対応付け技術が確立されました。
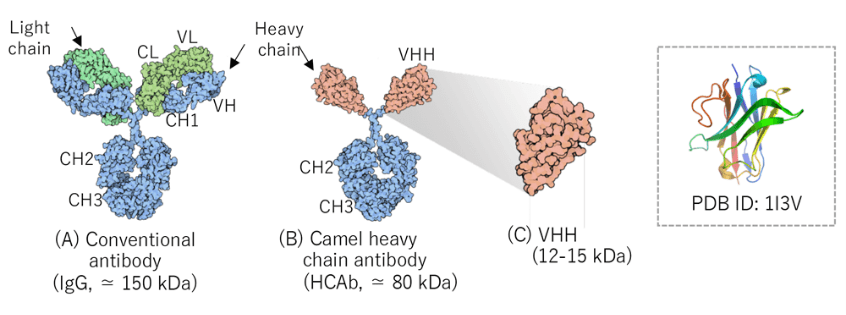
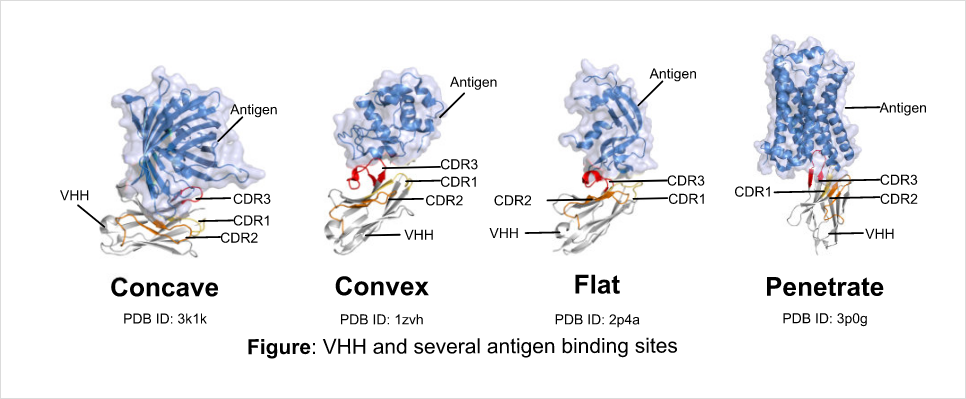
一方、VHHは1993年にベルギーのブリュッセル自由大学のHamers教授によって発見されました(Hamers-Casterman, C., et al. 1993))。創薬に有利な多くの特徴を有しているものの、抗体医薬品が次世代バイオ医薬品として注目を集め始めた時期と重なり、大企業から注目されることはありませんでした。しかし、ベルギーでVHHの医薬品化を目指すバイオベンチャー企業が立ち上がり、低分子化抗体医品の一つとして研究開発がすすめられました。そして、VHH抗体医薬の第1号として、2018年FDAは後天性血栓性血小板減少性紫斑病(aTTP)を適応に抗von-Willebrand因子に対するVHHを承認しました。これに続くかのようにVHH抗体医薬が続々と臨床フェーズへ進みつつあり、VHHは次世代バイオ分子としてさらに注目を浴びています。